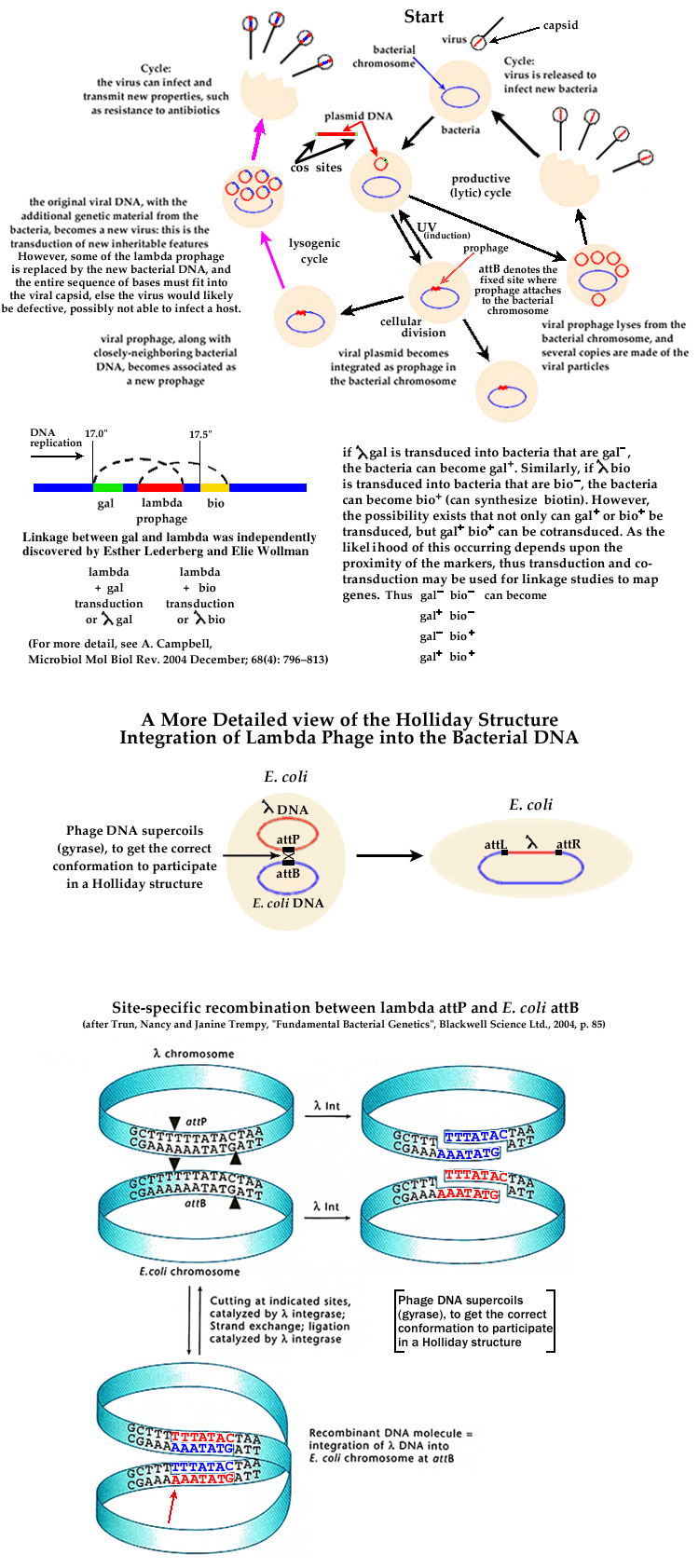
As the relationship between the molecular structure of DNA and genetic properties became a possibility, lambda bacteriophage became of immediate importance. One of the first areas of lambda research was in transduction: some bacteria and lambda phage led to the distinction between HFT (High Frequency Transducers) and LFT (Low Frequency Transducers).
See "The Bacteriophage Lambda" by A. D. Hershey, 1971, Cold Spring Harbor Press.
.
.
.
.
.
.
.
.
.
Possible Transduction Crosses:
Thus using transduction and cotransduction, it is possible to
create gene maps based upon linkage (A. D. Hershey and R. Rotman).
.
.
"While Zinder was pushing forward the work on Salmonella
transduction, Lederberg's laboratory was continuing work on all
aspects of genetics in Escherichia coli K-12. As discussed
in Section 7.5, phage lambda was discovered in this strain
by Esther Lederberg in 1951, and its
linkage to the gal locus was shown by mating (Lederberg and
Lederberg, 1953; Wollman, 1953). M. L. Morse, a student of
Lederberg, attempted to determine if lambda was capable
of transduction. Of numerous genetic markers tested, only a cluster
of genes for galctose fermentation were transduced by lambda
lysates (Morse, Lederberg, and Lederberg, 1956a). Also, transducing
lysates could only be obtained by induction of lysogenic
strains; lytic infection by free lambda phage particles
did not result in transducing lysates. Since lambda prophage
was linked to gal (see Section 7.5), the restriction of
transduction to the gal region suggested a different and more
special situation in lambda transduction than that for P22
in Salmonella. Because lambda transduction was
restricted to only a few genes, it came to be called specialized
transduction."
"The Emergence of Bacterial Genetics," Thomas D. Brock, Cold Spring
Harbor Laboratory Press, Cold Spring Harbor, NY, 1990, pp. 202-203.
It should be noted that the book above presents a biased view in favour of Joshua Lederberg: "...the work of Joshua Lederberg..., whom Brock has chosen as the hero of his story." See the review of the above book by Gunther S. Stent, Nature, 39: 661-662, Feb. 1991.
The importance of specialized transduction in lambda was commented upon by Norton Zinder himself in Current Contents 20, May 16, 1983 (p. 30). In this article he referred to Zinder, N.D. and Lederberg, J., "Genetic exchange in Salmonella," J. Bacteriol. 64: 679-699, 1952. Summarizing part of the article, Norton Zinder refers to genetic transduction, and notes that "we knew nothing about phage or temperate phage at the time." With reference to genetic transduction (not specialized transduction), he further points out that:
"General transduction3 provides the means for a fine genetic analysis of genome structure. The discovery shortly thereafter of special transduction4 complemented [general transduction] ... the paper also describes phage P22 (then called PLT-22), which became, with lambda, special transduction, the two paradigms for the study of temperate bacteriophages."
To get an idea of how much research was accomplished by the collaboration of Esther M. Zimmer Lederberg and M. Laurence Morse, examine the Galactosemia correspondence displayed at this website. Much of the correspondence, indeed, is focused upon gene mapping based upon transduction. Almost all the research on the relationshp between lambda and gal transduction was personally performed by Esther M. Lederberg and M. Laurance Morse, which is clearly shown in the correspondence (Joshua Lederberg was not involved in experiments). This is not surprising, since both Esther M. Lederberg and M. Laurance Morse were excellent experimentalists and, as other research scientists (such as Stan Falkow and Joshua Lederberg himself) have pointed out, Joshua Lederberg was not gifted with this ability. 5
As a note, the Fertility Factor F (the first episome discovered, and discovered by Esther M. Lederberg) is also capable of transduction. In "Episomes" by Allan Campbell (Harper & Row, NY, 1969), on page 105:
"The F episome resembles phage lambda in occasional occurrence of variants that have picked up genes from the bacterial chromosome. As with lambda, such variants are produced only from integrated F (i.e., from Hfr strains) and seem to comprise continuous blocks of genes contiguous to the F attachment site of the particular Hfr strain used."
Thus the lambda phage discovered by Esther M. Zimmer Lederberg was of major importance in explaining transduction. For a list of related papers, click here.
1 Multiple plasmids (if compatible) may be found in one prokaryote cell. In some cases, the different plasmids may lysogenize, forming several prophages simultaneously in the same prokaryote chromosome. As an alternative to the process described here for lambda phage, Mu (for "mutations") is another bacteriophage that works quite differently than lambda does. The Mu virus injects its linear DNA into the bacterial host (E. coli) cytoplasm. The linearized phage DNA remains linear in the host cytoplasm (subject to degradation) and integrates into the host DNA as a "prophage". Now, by replicative transposition (θ structures, or Shapiro intermediates), multiple copies of the Mu prophage are created in the host DNA (directly, not in the cytoplasm)! When excision takes place, multiple copies of the Mu DNA are then assembled into capsids, tails are assembled, and heads and tails are joined. The host cell bursts (using proteins that attack the host cell wall just as lambda does), releasing the multiple copies of Mu virus into the environment to further infect new host bacteria.
2 Lambda forms its prophage at a specific location in the E. coli chromosome (at site "attP" in the prophage [250 base pairs], "attB" is the matching bacterial DNA site [21 base pairs]). Thus only a few closely neighboring markers may participate in specialized transduction. Although taking place with far less probability, if "attB" is defective or absent, alternate sites with similar base sequences may participate in lysogeny, and thus different genes may be specially transduced. Other viral plasmids are not so restricted to specific bacterial sites, and as their prophages are not restricted to a specific location, these viral prophages can have many closely neighboring markers. This is generalized transduction. For this reason, lambda transduction is classified as specialized transduction and led to most of the detailed knowledge about how transducion actually functions.
3 "Transducing fragments in generalized transduction by phage P1", Ikeda, H. & Tomizawa, J., J. Molec. Biol. 1965, 14(1), 85-109
4 "Transduction in E. coli K-12", Morse, M., Lederberg, E. & Lederberg, J., 1956, Genetics, 41(1), 142-156
5 Thus major contributions to understanding specialized transduction and transduction in general came from the experiments by Esther M. Lederberg, Larry Morse, Joshua Lederberg, Norton Zinder, Allan Campbell, Werner Arber, etc. Citations are not provided as the number of relevant papers in this area from 1951 through 1956 and after is so large. In addition to the extensive correspondence with Larry Morse concerning specialized transductions with galactosemia, there also is an extensive correspondence concerning maltophilia research between Esther M. Lederberg and researchers such as Julius Adler, Enrico Calef, and Franco Guerrini (among others) concerning transduction and defective lambda.
Explanations at greater depth involving topics discussed here may
be found in the following two sources:
.
Related papers authored or co-authored by Esther M. Zimmer Lederberg:
Lederberg, E., 1950-1951, "Lysogenicity and Transduction E. coli",
referenced in 1957, "Symposium on Bacterial and Viral Genetics (Canberra,
August 1957)", p. 71
Lederberg, J., Lederberg, E. M., Zinder, N. D., Lively, E. R., 1951,
"Recombination analysis of bacterial heredity", Cold Spring Harbor
Symposia on Quantitative Biology 16:413-443
Morse, M. L., Lederberg, E. M., Lederberg, J., 1956, "Transduction
in Escherichia coli K-12", Genetics 41:142-156
Morse, M. L., Lederberg, E. M., Lederberg, J., Sept. 1956, "Transductional
heterogenotes in Escherichia coli", Genetics 41(5):758-779
© Copyright 2006 - 2018
The Esther M. Zimmer Lederberg Trust
 Web Site Terms of Use
Web Site Terms of Use